La fotosíntesis es un proceso físico-químico por el cual plantas, algas, bacterias fotosintéticas y algunos protistas como diatomeas utilizan la energía de la luz solar para sintetizar compuestos orgánicos. Se trata de un proceso fundamental para la vida sobre la tierra y tiene un profundo impacto sobre la atmósfera y el clima terrestres: cada año los organismos con capacidad fotosintética convierten en carbohidratos más del 10% del dióxido de carbono atmosférico. El conocimiento básico de este proceso es esencial para entender las relaciones entre los seres vivos y la atmósfera así como el balance de la vida sobre la tierra.

En plantas, algas y en algunos tipos de bacterias fotosintéticas el proceso conlleva la liberación de oxígeno molecular y la utilización de dióxido de carbono atmosférico para la síntesis de compuestos orgánicos. A este proceso se le denomina fotosíntesis oxigénica.
Sin embargo, algunos tipos de bacterias utilizan la energía de la luz para formar compuestos orgánicos pero no producen oxígeno. En este caso se habla de fotosíntesis anoxigénica.
Antes de existir oxígeno en la atmósfera, el ambiente de las primeras formas de vida era anaerobio. Estos primeros organismos no tenían capacidad para sintetizar sus propios nutrientes orgánicos y tomaban del medio lo que ya estaba sintetizado. Eran heterótrofos. Estos heterótrofos primitivos seguían alimentándose del medio, pero el medio iba cambiando: la tierra se iba enfriando, iba disminuyendo la radiación ultravioleta que alcanzaba la superficie terrestre, etc. Y en este escenario se produjo un cambio que consistió en ser capaz de sintetizar las moléculas energéticas. Entonces los organismos se hacen autótrofos. En todo caso, estamos hablando de nutrición, es decir, de los componentes necesarios para la supervivencia, o lo que es lo mismo, de fuentes de carbono, nitrógeno, hidrógeno y energía. Y dependiendo de cuáles son estas fuentes, denominamos a los distintos organismos.

Para que el proceso fotosintético ocurra, para que se inicie la fase fotoquímica (conversión de la energía de la luz en energía química), lo primero que tienen que hacer los organismos es captar la luz. Las moléculas que intervienen en ello son los pigmentos fotosintéticos, los cuales se organizan, se colocan, en una membrana: la membrana plasmática en bacterias, y la membrana tilacoidal de los cloroplastos de cianobacterias, algas y plantas.
CAPTACIÓN DE LUZ Y TRANSPORTE FOTOSINTÉTICO DE ELECTRONES
Todos los organismos con capacidad fotosintética contienen uno o más pigmentos capaces de absorber radiación visible que desencadena las reacciones fotoquímicas de la fotosíntesis. Estos pigmentos se pueden extraer de los organismos que los contienen con alcohol o con disolventes orgánicos (recordemos su naturaleza química).
Parte de la energía luminosa absorbida por clorofilas y carotenoides se almacena al final del proceso fotosintético como energía química. La mayoría de los pigmentos actúan como una antena (en un complejo antena) captando la luz y transfiriendo la energía (proceso físico) al centro de reacción al que están asociados y donde se transfieren electrones desde la clorofila a una molécula aceptora de electrones (proceso químico).

FOTOSISTEMA I Y FOTOSISTEMA II
Existen dos complejos fotoquímicos denominados fotosistema I (PSI) y fotosistema II (PSII) en los que tienen lugar las reacciones iniciales de almacenamiento de energía. Si ambos fotosistemas funcionan en serie se producen dos reacciones fotoquímicas en serie. El PSI absorbe luz del rojo lejano de 700nm (longitudes de onda superiores a 680 nm), produce un reductor fuerte capaz de reducir NADP+ y un oxidante débil. El PSII absorbe luz del rojo de 680nm, produce un oxidante muy fuerte capaz de oxidar al agua y un reductor más débil que el producido por el PSI. La figura 6 muestra un esquema de estas propiedades que se conoce como esquema en Z, implica ambos fotosistemas y explica las reacciones fotoquímicas que ocurren en los organismos fotosintéticos que generan oxígeno (recordemos fotosíntesis oxigénica y fotosíntesis anoxigénica). Ambos fotosistemas son física y químicamente diferentes, contienen cada uno su complejo antena y su centro de reacción, y están unidos por una cadena de transporte electrónico.

Además de PSII y PSI, englobados en la membrana tilacoidal se encuentran también otros dos complejos proteicos: el complejo citocromo b6f y la ATP sintasa.
La clorofila de los centros de reacción, como ya sabemos, es una clorofila especializada que denominamos P680 en el PSII y P700 en el PSI (haciendo referencia a sus características espectrales). El funcionamiento básico de todo este conjunto sería el siguiente:
• El PSII oxida el agua y produce O2 liberando protones al lumen tilacoidal
• El complejo cit b6f recibe electrones del PSII y los cede al PSI. También transporta protones al lumen desde el estroma
• El PSI reduce el NAP+ a NADPH en el estroma gracias a la acción de una ferredoxina (fd) y una flavoproteína ferredoxina-NADP reductasa (FNR)
• La ATP sintasa produce ATP en el estroma a medida que los protones difunden a su través desde el lumen hacia el estroma.
FASES
Fase luminosa.
Fase en donde se transforma la energía luminosa en química:
que es usada por todos los seres vivos. Los vegetales son el primer y único
eslabón productor de la cadena trófica.
Esta fase depende de la luz que reciben los cloroplastos de la células vegetales
que son captados por medio de la clorofila, esta energía lumínica descompone el
agua en Oxigeno e Hidrogeno, liberándose el Oxigeno y generándose 2
moléculas por medio del movimiento de sus electrones de un nivel a otro liberando
energía para producir la molécula ATP y el poder reductor que es la molécula
NADPH2 que aportaran a la fase siguiente energía química para la transformación
de CO2 en Hidratos de carbono.

Fase Oscura.
En la Fase oscura en la que ya no interviene la luz y las moléculas formadas en
la fase luminosa (ATP y NADPH2) participan en la reducción del bióxido de
carbono (CO2) mediante una serie de reacciones el “Ciclo de Calvin” en donde se
combina Se combina CO2 con RDP (difosfato de ribulosa) para formar PGA (ác.
Fosfoglicérido)
Se combina PGA con NADPH2 y ATP por lo que se libera agua, se forma PGAL
para la nutrición de la planta, se produce glucosa a partir de PGAL, este azúcar
se disuelve en agua y recorre toda la planta proporcionándole la energía necesaria
para crecer.
Mecanismo C4, plantas C4
Existen diferencias anatómicas en las hojas de plantas C4. La siguiente figura muestra un esquema de la sección transversal de la hoja.

Como podemos observar, en las hojas de plantas C4 hay dos clases de células con cloroplastos, las células del mesófilo y las células de la vaina del haz que se ordenan de forma distinta a las del mesófilo. Además las del mesófilo están próximas a la de la vaina y hay una extensa red de plasmodesmos que las conecta facilitando todo ello la cooperación fisiológica, el flujo de metabolitos, entre ambos tipos de células.
El carácter distintivo de las plantas C4 es la fijación inicial de CO2 en un ácido dicarboxílico de 4 átomos de C, el ácido oxalacético, en el citosol de la célula del mesófilo, en una reacción de carboxilación de fosfoenolpirúvico catalizada por la enzima fosfoenolpirúvico carboxilasa. Este es el inicio de las cuatro etapas que completan el ciclo C4 en las células del mesófilo y del haz de la vaina. Las etapas que se esquematizan en la siguiente figura son:
1) fijación de C en un ácido de 4C en el mesófilo.
2) Transporte de ácidos 4C desde el mesófilo a las células de la vaina.
3) descarboxilación de los ácidos 4C generándose una alta concentración de CO2 en células de la vaina. Este CO2 es fijado por la rubisco dirigiéndose al ciclo de Calvin.
4) Transporte del ácido de 3C resultante a la célula del mesófilo donde se regenera el aceptor inicial, fosfoenolpiruvato.
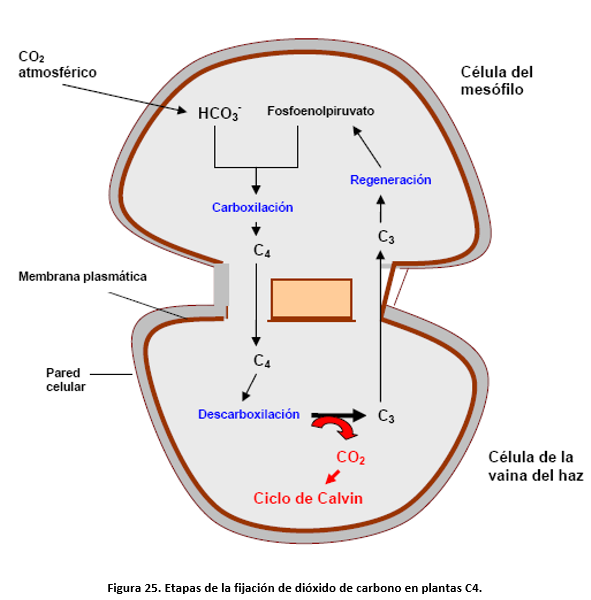
Este ciclo transporta de forma muy efectiva CO2 desde la atmósfera hasta las células de la vaina, produciéndose en estas una concentración tal de CO2 que se inhibe la fotorrespiración. La figura 26 muestra un esquema más completo del ciclo C4 en el que se puede apreciar que la regeneración del aceptor requiere, consume, 2 ATP por molécula de CO2 transportada. En consecuencia, en una planta C4 la energía total requerida para fijar una molécula de CO2 es de 5 moléculas de ATP y 2 moléculas de NADPH.
Pero las plantas C4 difieren entre ellas en la metabolización del oxalacetato y en la etapa de descarboxilación. Se pueden distinguir tres tipos o grupos:
1) Plantas C4 NADP-ME: en las que el oxalacetato se reduce a malato, son formadoras de malato, que posteriormente es descarboxilado en el cloroplasto de las células de la vaina por la enzima málico-NADP dependiente (corresponde al esquema de la figura anterior).
2) Plantas C4 NAD-ME: en las que la enzima málico responsable de la descarboxilación es dependiente de NAD y mitocondrial.
3) Plantas C4 PEP-CK: en las que la descarboxilación está catalizada por la enzima fosfoenolpirúvico carboxiquinasa.
El ciclo C4 está regulado por luz de manera que la actividad de varios enzimas responden a variaciones en la densidad de flujo a través de oxido-reducción de grupos tiol y fosforilación-defosforilación.
Las plantas C4, la fotosíntesis C4 podría decirse, es una adaptación evolutiva a condiciones ambientales de alta irradiancia y temperatura, y ciertas limitaciones de agua nunca severas, condiciones propias de regiones tropicales y templadas. Entre ellas se incluyen plantas de gran importancia económica e industrial como el maíz, la caña de azúcar o el sorgo, entre las especies cultivadas. También muchas gramíneas, la mayor parte “malas hierbas” de rápido crecimiento y gran capacidad de competencia con otras especies.
Mecanismo CAM, plantas CAM
El mecanismo CAM de concentración de CO2 se describió inicialmente en plantas crasuláceas de donde toma el nombre. Pero no está restringido a esta familia sino que se encuentra en otras muchas, por ejemplo, cactus, euforbiaceas, piña, vainilla, etc. Las plantas CAM presentan un proceso diario durante la noche de acumulación de ácidos orgánicos, ácido málico fundamentalmente, que después degradan durante el día. Son plantas adaptadas a condiciones de aridez extrema de manera que abren los estomas durante la noche, para minimizar la pérdida de agua por transpiración, fijando CO2 en oscuridad (por la noche) por una reacción de carboxilación de PEP (fosfoenolpirúvico) catalizada por PEP carboxilasa en el citosol. El resultado es la formación de oxalacetato y malato que se almacena en la vacuola resultando de ello una acidificación nocturna de la hoja .

El malato almacenado en la vacuola se libera durante el día, se transporta al cloroplasto, cuando los estomas están cerrados. En el cloroplasto el malato es descarboxilado por la enzima málico NADP dependiente y el CO2 liberado se fija en el ciclo de Calvin. El ácido pirúvico es convertido de nuevo en azúcares y finalmente en almidón.
En el proceso se acumulan de forma transitoria malato y almidón. Este último, el almidón, actúa como precursor de fosfoenolpiruvato, el aceptor del CO2, que a diferencia de las C4 no se regenera continuamente.
La actividad de la PEP carboxilasa es baja durante el día y alta durante la noche. El mecanismo implica la separación de la carboxilación inicial de la descarboxilación posterior para evitar ciclos inútiles. Pero la regulación de la PEP carboxilasa es clave para la eficacia del proceso. Esta enzima es inhibida por malato y activada por glucosa-6-fosfato pero la fosforilación de un residuo de serina de la enzima de las plantas CAM disminuye la inhibición por malato y aumenta la acción de glucosa-6fosfato. Esta fosforilación de fosfoenolpirúvico carboxilasa está catalizada por PEP carboxilasa quinasa.

El mecanismo CAM está asociado en muchas especies a la suculencia, una característica que confiere a las plantas una baja relación superficie/volumen que limita el intercambio de gases y aumenta su capacidad de almacenamiento.
Pero algunas plantas son capaces de adaptar su metabolismo a las condiciones ambientales de manera que pueden presentar un ciclo CAM de carácter adaptativo. Estas plantas se comportan como C3 induciéndose el ciclo CAM bajo determinadas circunstancias. Por ello se les denomina CAM facultativas. Por ejemplo, Mesembryanthemum crystallinum realiza ciclo C3 en condiciones normales de no estrés, pero cambia a ciclo CAM es respuesta a estrés por calor, salino o hídrico.
En ambientes acuáticos, las cianobacterias y las algas tienen agua abundante pero menor disponibilidad, por haber menor concentración, de CO2 por lo que, como vimos al principio, concentran activamente CO2 intracelular.









